High-quality SNPs from genic regions highlight introgression patterns among European white oaks (Quercus petraea and Q. robur)
Tiange Lang, Pierre Abadie, Valérie Léger, Thibaut Decourcelle, Jean-Marc Frigerio, Christian Burban, Catherine Bodénès, Erwan Guichoux, Grégoire Le Provost, Cécile Robin, Naoki Tani, Patrick Léger, Camille Lepoittevin, Veronica A. El Mujtar, François Hubert, Josquin Tibbits, Jorge Paiva, Alain Franc, Frédéric Raspail, Stéphanie Mariette, Marie-Pierre Reviron, Christophe Plomion, Antoine Kremer, Marie-Laure Desprez-Loustau, Pauline Garnier-Géré
https://doi.org/10.1101/388447
A new resource for a better understanding of the complex genetic dynamics within the oak syngameon
Recommended by Erwin Dreyer based on reviews by Oliver Brendel, Ricardo Alia, Komlan Avia and Hilke Schröder based on reviews by Oliver Brendel, Ricardo Alia, Komlan Avia and Hilke Schröder
When we launched the multidisciplinary PCI Forest and Wood Sciences, we were well aware that this PCI devoted to a broad object (forests represent 1/3 of the emerged land area) and gathering multiple disciplines (from biology to social sciences), would be confronted with some challenges with respect to the range of pre-prints we would have to consider. Among the first ones submitted to the new PCI was this very interesting one (Lang et al, 2021) in the area of tree genomics and tree population genetics with a strong background in bioinformatics. This could well have been submitted to the PCI in Genomics, or the PCI in Evolutionary Biology, but finally fits very well in PCI Forest and Wood Sciences due to its finality: provide reliable genomic resources to tackle questions that are central to our understanding of the dynamics of forest ecosystems and the adaptation of trees to local conditions.
The phylogenetic status of the two sympatric oak species Quercus robur L. and Q. petraea Matt. Liebl. is an important question that was the center of in depth investigations by the BioGeCo research group at Bordeaux in cooperation with many European groups. The two species are major forest tree species in western Europe, structuring important ecosystems and providing high quality wood and other important ecosystem services. These species are considered to build up what is now called a “syngameon” together with a few other western European white oaks (Cannon and Petit, 2019). A syngameon is a complex of species exchanging genes, while still maintaining some degree of interspecific diversity enabling to differentiate, on the basis of leaf morphology for instance, the originally described species (Kremer et al, 2002 among many others dealing with the identification of oak species on the basis of their phenotype). The full sequencing of the genome of Q. robur (Plomion et al, 2018) opened an avenue for the development of new genomic tools to investigate the concept of species in this syngameon and to identify gene flows between populations and species. To date, whole genome sequencing and annotation is completed for Q. robur, Q. lobata Née, Q. suber L., and Fagus sylvatica L. (Plomion and Martin, 2020).
The preprint of Lang et al (2021) builds up on this background and provides a very interesting and important contribution to this area. First, it provides access to a very large data base of genomic sequences, based on 800 gene fragments and identified 12500 Single Nucleotide Polymorphisms (SNPs) as well as 1500 insertions deletions. 11 individuals assigned to the species Q. robur, 13 assigned to Q. petraea and 1 to Q. ilex L. were used to establish this data base. This is an important asset for developing future research on species identification based on their genomes. In full agreement to the policy of all PCIs about data availability, full access is provided to the data under a free use license CC-BY.
The manuscript has already a long history as a preprint and it was not that easy to find reviewers willing to do the job of reviewing such a large amount of information with a lot of technical details. At the end, we succeeded and obtained three very enthusiastic and positive reviews (attached to this recommendation). All reviewers agreed that this was an important contribution in the field, insisted on the quality of the writing and of the provided data, and finally agreed that the manuscript was to be recommended after a careful revision. The revision was readily done by the authors. One of the reviewers found that the preprint is close to a “data paper”, i.e., a paper providing the required information to enable (and advertise) the reuse of an important data set, which is only partly true. Indeed, the authors nicely used their data to address one of the most debated questions about the population genetics: how comes that in this syngameon, with inter-fertile individuals, clearly distinguishable species (from the phenotype at least) have been maintained?
The main result of the preprint is that indeed there are some (only few) genomic differences among the two species in the patterns of distribution of the SNPs, but also that the patterns reveal a clear introgression of the genome of one of the species into the other one. This was based on a Bayesian clustering analysis reported figure 3, evidencing that 2 individuals displaying an unambiguous Q. petraea phenotype could not be assigned to any of the species. The usual hypothesis postulates an introgression of the Q. robur genome, the pioneer species able to colonize open areas during the post-glacial recolonization into that of Q. petraea, due to hybridization and successive events of back gross with inflowing Q. petraea pollen. Such an introgression was hypothesized decades ago, and receives here some degree of confirmation. Recent findings showed the importance of such an introgression for the adaptation of local oak populations to altitudinal gradients (Leroy et al, 2020).
Given the importance of the contribution (both from the point of view of the development of new genomic tools and from that of our understanding of the population genetics in the oak syngameon, I do recommend this preprint in the frame of the PCI Forest and Wood Sciences after an insightful peer reviewing and a very careful revision leading to the current version 4 of this preprint.
References
Cannon CH, Petit RJ (2020) The oak syngameon: more than the sum of its parts. New Phytologist, 226, 978–983. https://doi.org/10.1111/nph.16091
Kremer A, Dupouey JL, Deans JD, Cottrell J, Csaikl U, Finkeldey R, Espinel S, Jensen J, Kleinschmit J, Dam BV, Ducousso A, Forrest I, Heredia UL de, Lowe AJ, Tutkova M, Munro RC, Steinhoff S, Badeau V (2002) Leaf morphological differentiation between Quercus robur and Quercus petraea is stable across western European mixed oak stands. Annals of Forest Science, 59, 777–787. https://doi.org/10.1051/forest:2002065
Lang T, Abadie P, Léger V, Decourcelle T, Frigerio J-M, Burban C, Bodénès C, Guichoux E, Provost GL, Robin C, Tani N, Léger P, Lepoittevin C, Mujtar VAE, Hubert F, Tibbits J, Paiva J, Franc A, Raspail F, Mariette S, Reviron M-P, Plomion C, Kremer A, Desprez-Loustau M-L, Garnier-Géré P (2021) High-quality SNPs from genic regions highlight introgression patterns among European white oaks (Quercus petraea and Q. robur). bioRxiv, 388447, version 4 peer-reviewed and recommended by PCI Forest and Wood Sciences. https://doi.org/10.1101/388447
Leroy T, Louvet J-M, Lalanne C, Provost GL, Labadie K, Aury J-M, Delzon S, Plomion C, Kremer A (2020) Adaptive introgression as a driver of local adaptation to climate in European white oaks. New Phytologist, 226, 1171–1182. https://doi.org/10.1111/nph.16095
Plomion C, Aury J-M, Amselem J, Leroy T, Murat F, Duplessis S, Faye S, Francillonne N, Labadie K, Le Provost G, Lesur I, Bartholomé J, Faivre-Rampant P, Kohler A, Leplé J-C, Chantret N, Chen J, Diévart A, Alaeitabar T, Barbe V, Belser C, Bergès H, Bodénès C, Bogeat-Triboulot M-B, Bouffaud M-L, Brachi B, Chancerel E, Cohen D, Couloux A, Da Silva C, Dossat C, Ehrenmann F, Gaspin C, Grima-Pettenati J, Guichoux E, Hecker A, Herrmann S, Hugueney P, Hummel I, Klopp C, Lalanne C, Lascoux M, Lasserre E, Lemainque A, Desprez-Loustau M-L, Luyten I, Madoui M-A, Mangenot S, Marchal C, Maumus F, Mercier J, Michotey C, Panaud O, Picault N, Rouhier N, Rué O, Rustenholz C, Salin F, Soler M, Tarkka M, Velt A, Zanne AE, Martin F, Wincker P, Quesneville H, Kremer A, Salse J (2018) Oak genome reveals facets of long lifespan. Nature Plants, 4, 440–452. https://doi.org/10.1038/s41477-018-0172-3
Plomion C, Martin F (2020) Oak genomics is proving its worth. New Phytologist, 226, 943–946. https://doi.org/10.1111/nph.16560
| High-quality SNPs from genic regions highlight introgression patterns among European white oaks (Quercus petraea and Q. robur) | Tiange Lang, Pierre Abadie, Valérie Léger, Thibaut Decourcelle, Jean-Marc Frigerio, Christian Burban, Catherine Bodénès, Erwan Guichoux, Grégoire Le Provost, Cécile Robin, Naoki Tani, Patrick Léger, Camille Lepoittevin, Veronica A. El Mujtar, Fran... | <p>In the post-genomics era, non-model species like most Fagaceae still lack operational diversity resources for population genomics studies. Sequence data were produced from over 800 gene fragments covering ~530 kb across the genic partition of E... |  | Population dynamics, genetics and genomics of forest trees | Erwin Dreyer | | 2020-03-03 08:00:45 | View |
A very useful simulation of the impact of drought and enhanced temperatures on embolism in trees and on tree dieback
Recommended by Erwin Dreyer based on reviews by Sabine Rosner and 1 anonymous reviewer
Water availability has been known to strongly modulate forest productivity and tree growth on an interannual basis (as revealed by numerous dendrochronological studies) and across biomes (Ellison et al, 2017). Recurrent episodes of severe drought lead to decreased soil water content and as a consequence to visible losses in annual growth increment, and in some cases even to tree death and forest decline. The occurrence of such drought events and of larger scale tree dieback, seem to be increasing over the last decades, albeit such processes are not new. The causes for drought-induced tree death are still disputed; in many cases, tree death occurs after the release of drought, and is caused by severe attacks by pests and pathogens. In other cases, tree death is caused by recurrent drought events over several years, leading to a depletion of stored carbohydrates, growth decline and ultimately death.
However, this understanding of drought-induced tree dieback, which applies to drought events that occurred in temperate climate biomes during the end of the 20th century, seems inadequate to explain the increasing occurrence of large scale dieback induces by recent drought episodes (Allen et al, 2015). In these recent cases a direct impairment of hydraulic functions seems responsible for tree death. Such impairments (cavitation and resulting massive embolism) have been well documented through extensive research that started in the 90s. Up to now, the consensus was that trees are fairly well protected against such potentially lethal dysfunctions: an efficient stomatal closure limits transpiration and the risk of runaway embolism. Many tree models based on the known hydraulic properties of trees (vulnerability of different organs to cavitation, hydraulic conductance of these organs, transpiration, stomatal conductance…) were developed since the seminal work of Tyree and Sperry (1989) and only seldom predicted the occurrence of runaway embolism (Cochard and Delzon, 2013).
These models considered the impact of drought through reduced soil water availability, which is indeed the central process during drought, but overlooked to some extent the fact that drought is frequently and increasingly associated to higher temperatures, which may change rather severely model parameters and result in a higher risk of runaway embolism.
The present preprint proposed by Cochard (2020) bases on such a new hydraulic model (the model SurEau, Martin StPaul et al, 2017; Cochard et al, 2020) integrating more explicitly the impact of temperature on different parameters. Two parameters appear particularly relevant and highly sensitive to temperature:
(i) the vapor pressure deficit of the air (VPD), which increases exponentially with temperature and results in increased transpiration and more rapid soil water depletion; this effect is well known and has been the matter of many research and modelling;
(ii) the cuticular conductance to water vapor, which becomes the most important limit to transpiration once stomata are closed, and which is much less well documented with respect to mean values and temperature sensitivity (mainly because this process is difficult to record). Recent advances (Schuster et al, 2016) provided some insight into the importance of this parameter and showed how it may rapidly increase with temperature (see references in the preprint).
The presented work bases on this new model to document more precisely how enhanced temperature may increase water loss through transpiration and consequently induce runaway embolism in trees more rapidly than usually expected. The hypothesis that the temperature response of cuticular conductance may play a central role in the sensitivity of trees to a combination of soil water depletion and enhanced air (and leaf) temperature was tested through numerical simulations with SurEau. The results are very clear: temperature-dependent increases in cuticular conductance may accelerate the onset of runaway embolism at a rate that was not expected before.
The demonstration is indeed very clear and convincing. It remains however a simulation (or an “in silico experiment”. Data providing real values of cuticular conductance remain scarce, and data documenting its response to enhanced temperatures even scarcer. This opens an avenue for new research and investigations, and Cochard (2020) provides some clues about which data and which experiments could confirm the central role of temperature induced changes in cuticular conductance with temperature (eg new measurements of Tp, the phase transition temperature that matches the range of temperatures known to trigger mortality during hot-droughts, Billon et al. (2020)).
I believe this preprint is an important contribution in this field, and the reviewers were of the same opinion (see their reviews attached to this recommendation). Indeed, this preprint illustrates how simulation exercises can help us identify some key processes that require further attention and documentation. I believe this is an important contribution to our understanding of the rapid, drought-induced tree death observed in different parts of the world at alarming rates.
As such, and combined with a detailed description of the model SurEau, this preprint is a very important addendum to the corpus of knowledge that is currently gathered around the hydraulic functioning of trees.
References
Allen, C. D., Breshears, D. D., and McDowell, N. G. (2015). On underestimation of global vulnerability to tree mortality and forest die‐off from hotter drought in the Anthropocene. Ecosphere, 6(8), 1-55. doi: https://doi.org/10.1890/ES15-00203.1
Billon et al. (2020). The DroughtBox: A new tool for phenotyping residual branch conductance and its temperature dependence during drought. Plant, Cell and Environment, 43, 1584-1594. doi https://doi.org/10.1111/pce.13750
Cochard, H. (2020) A new mechanism for tree mortality due to drought and heatwaves. bioRxiv, 531632, ver. 2 peer-reviewed and recommended by PCI Forest and Wood Sciences. doi: https://doi.org/10.1101/531632
Cochard, H., Martin-StPaul, N., Pimont, F., and Ruffault, J. (2020). SurEau.c: a mechanistic model of plant water relations under extreme drought. bioRxiv, 2020.05.10.086678. doi: https://doi.org/10.1101/2020.05.10.086678
Ellison et al. (2017). Trees, forests and water: Cool insights for a hot world. Global Environmental Change, 43, 51-61. doi: https://doi.org/10.1016/j.gloenvcha.2017.01.002
Martin‐StPaul, N., Delzon, S., and Cochard, H. (2017). Plant resistance to drought depends on timely stomatal closure. Ecology letters, 20(11), 1437-1447. doi: https://doi.org/10.1111/ele.12851
Schuster et al. (2016). Effectiveness of cuticular transpiration barriers in a desert plant at controlling water loss at high temperatures. AoB Plants, 8(1), plw027. doi: https://doi.org/10.1093/aobpla/plw027
Tyree, M. T., and Sperry, J. S. (1989). Vulnerability of xylem to cavitation and embolism. Annual review of plant biology, 40(1), 19-36. doi: https://doi.org/10.1146/annurev.pp.40.060189.000315
| A new mechanism for tree mortality due to drought and heatwaves | Hervé Cochard | <p>Plants tend to die earlier in hot and drought conditions, but the underlying mechanisms are not yet understood. I propose here a new mechanism by which excessive residual water losses caused by high cuticular permeabilities and a high leaf-to-a... | 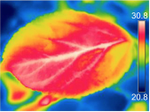 | Tree biology and physiology | Erwin Dreyer | | 2020-03-03 09:33:12 | View |
Interaction of drought and frost in tree ecophysiology: rethinking the timing of risks
Guillaume Charrier, Nicolas Martin-Stpaul, Claire Damesin, Nicolas Delpierre, Heikki Hänninen, José Torres-Ruiz, Hendrik Davi
https://hal.archives-ouvertes.fr/hal-02475505v4
Giving a temporal context to drought and frost vulnerability of trees
Recommended by Rosana López based on reviews by Jordi Martínez Vilalta and Sean Gleason
Range limits of forest species are frequently imposed by spatial gradients in climatic variables. Tolerance to maximum and minimum temperatures, including short-term extremes, and tolerance to desiccation are crucial limiting factors for plant survival and often appear interrelated (Box 1995; Choat et al. 2018). Increasing temperatures, more frequent and extreme droughts and late frost events associated with global climate change will affect the dynamics of forest ecosystems and have the potential to dramatically increase plant mortality and accelerate species range shifts if plants are not able to adapt in situ to the novel climate regimes (Parmesan 2006; Choat et al., 2012). This is particularly important at species range edges, where climatic conditions may not be sufficient to impose mortality on individuals directly, but trees experience greater physiological stress, which influences such factors as dispersal, habitat selection, and subsequent reproductive fitness (Parmesan 2006). In such marginal situations, where gene flow may be also restricted (López de Heredia et al. 2010), the effectiveness of adjustment through natural selection is limited resulting in increased vulnerability to extreme climatic events and to a higher risk of mortality of trees.
Tree responses to drought and frost have been extensively studied at many scales from ecophysiology to molecular biology across a large range of species inhabiting diverse biomes (Sakai and Larcher 1987; Bréda et al. 2006). Avoiding dehydration of tissues to maintain cellular viability and function is at the basis of the plant strategy to deal with both constraints as shown by Charrier et al. (2020). These authors go one step further and discuss the impact of the interaction of drought and frost on tree water status and carbon metabolism with special emphasis on the temporal context. Plants from temperate and boreal regions show changes in their resistance to freezing temperature throughout the year (Bower and Aitken 2006) and xylem becomes more resistant to cavitation with cambial age (Rodríguez-Zaccaro et al. 2019). Including timing in this framework involves incorporating phenology. This will be a fundamental step to model species distribution limits in the face of climate change since most observations of climate-change responses have involved alterations of species’ phenologies (Parmesan et al. 2006). For example, the onset of the growing season of trees in temperate Europe is 2.3 days ahead per decade during the last 40 years (Parmesan et al. 2006). Moreover, some studies have shown that climatic constraints limit species distribution mainly because of their impact in phenology rather than their impact on drought and frost mortality (Morin et al. 2007). However, longer growing seasons along with more frequent extreme events increase the probability of long-lived organisms such as trees to experience frosts and drought during the same growing season or one of them after uncompleted recovery of the other in some latitudes. Over the last decade much attention has been devoted to the recovery of growth and ecological function after stressful events (Lloret et al. 2011). Some dendroecological studies have shown for example that resilience to extreme droughts might be constrained by having experienced more frequent droughts, thus exceeding the potential for acclimation of the tree (Bose et al. 2020). Drought can result in chronic hydraulic impairment which can last moths to years (Anderegg et al. 2015) and for some species also in cavitation fatigue, i.e. a progressive increase in vulnerability to cavitation (Hacke et al. 2001), thus increasing vulnerability to subsequent drought and frost events.
Models have proved to be useful tools for synthetizing and integrating climate and soil properties with key functional traits in order to determine desiccation dynamics, carbon metabolism and plant survival during drought of frost (Martin-StPaul et al., 2017; Charrier et al. 2018; Blackman et al., 2019). However, models are currently limited by gaps in our understanding of the fundamental physiological mechanisms that constrain species ranges. Most common approaches to studying species range shifts are related to climatic niches and overlook the processes and traits involved in drought or frost tolerance (Cheaib et al. 2012). Process-based models based on plant hydraulics seem promising providing the link between environmental cues and plant responses, although disregarding carbon metabolism will not give us predictive understanding of system changes such as those due to climate fluctuations (Mackay et al. 2015). New modeling approaches need to be developed not only for better drought prediction performance but for the interaction of drought with other factors. Charrier et al. (2020) offer a framework for improving process-based models with the aim to provide better prediction of carbon and water economy, organ development and ultimately species distribution limits in the face of warmer winters and more frequent winter droughts at high altitudes and late frosts events.
References
Anderegg WR, Schwalm C, Biondi F, Camarero JJ, Koch G, Litvak M, Ogle K, Shaw JD, Shevliakova E, Williams A (2015) Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 349: 528–532. doi: https://doi.org/10.1126/science.aab1833
Blackman CJ, Li X, Choat B, Rymer PD, De Kauwe MG, Duursma RA, Tissue DT, Medlyn BE (2019) Desiccation time during drought is highly predictable across species of Eucalyptus from contrasting climates. New Phytologist 224: 632-643. doi: https://doi.org/10.1111/nph.16042
Box EO (1995) Factors determining distributions of tree species and plant functional types. Vegetatio 121, 101–116 (1995). doi: https://doi.org/10.1007/BF00044676
Bower AD, Aitken SN (2006) Geographic and seasonal variation in cold hardiness of whitebark pine. Can J For Res 36:1842–1850. doi: https://doi.org/10.1139/x06-067
Bréda N, Huc R, Granier A, Dreyer E (2006) Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 63: 625-644. doi: https://doi.org/10.1051/forest:2006042
Charrier G, Lacointe A, Améglio T (2018) Dynamic modeling of carbon metabolism during the dormant period accurately predicts the changes in frost hardiness in walnut trees Juglans regia L. Frontiers in Plant Science, 9: 1746. doi: https://doi.org/10.3389/fpls.2018.01746
Charrier G, Martin-Stpaul N, Damesin C, Delpierre N, Hänninen H, Torres-Ruiz J, Hendrik Davi H (2020) Interaction of drought and frost in tree ecophysiology: rethinking the timing of risks. HAL, 02475505, ver. 4 peer-reviewed and recommended by PCI Forest & Wood Sciences. https://hal.archives-ouvertes.fr/hal-02475505v4
Cheaib A, Badeau V, Boe J, Chuine I, Delire C, Dufrêne E, François C, Gritti ES, Legay M, Pagé C (2012) Climate change impacts on tree ranges: model intercomparison facilitates understanding and quantification of uncertainty. Ecology Letters 15(6): 533-544. doi: https://doi.org/10.1111/j.1461-0248.2012.01764.x
Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, et al. (2012) Global convergence in the vulnerability of forests to drought. Nature 491, 752–755. doi: https://doi.org/10.1038/nature11688
Choat B, Brodribb TJ, Brodersen CR, Duursma RA, López R, Medlyn BE (2018) Triggers of tree mortality under drought. Nature 558(7711): 531-539. doi: https://doi.org/10.1038/s41586-018-0240-x
Hacke UG, Stiller V, Sperry JS, Pittermann J, McCulloh KA (2001) Cavitation fatigue. Embolism and refilling cycles can weaken the cavitation resistance of xylem. Plant Physol., 125(2), 779–786. doi: https://doi.org/10.1104/pp.125.2.779
Lloret F, Keeling EG, Sala A (2011) Components of tree resilience: effects of successive low‐growth episodes in old ponderosa pine forests. Oikos, 120: 1909-1920. doi: https://doi.org/10.1111/j.1600-0706.2011.19372.x
López de Heredia U, Venturas M, López R, Gil L (2010) High biogeographical and evolutionary value of Canary Island pine populations out of the elevational pinebelt: the case of a relict coastal population. J. Biogeogr. 37, 2371–2383. doi: https://doi.org/10.1111/j.1365-2699.2010.02367.x
Mackay DS, Roberts DE, Ewers BE, Sperry JS, McDowell NG, Pockman WT (2015) Interdependence of chronic hydraulic dysfunction and canopy processes can improve integrated models of tree response to drought, Water Resour. Res., 51, 6156–6176. doi: https://doi.org/10.1002/ 2015WR017244
Martin-StPaul N, Delzon S, Cochard H (2017) Plant resistance to drought depends on timely stomatal closure. Ecology Letters 20(11): 1437-1447. doi: https://doi.org/10.1111/ele.12851
Morin X, Augspurger C, Chuine I (2007) Process-based modeling of species' distributions: what limits temperate tree species' range boundaries? Ecology 88(9):2280-2291. doi: https://doi.org/10.1890/06-1591.1 PMID: 17918406.
Parmesan C (2006) Ecological and evolutionary responses to recent climate change. Annual Review of Ecology and Systematics 37, 637–669. doi: https://doi.org/10.1146/annurev.ecolsys.37.091305.110100
Rodriguez‐Zaccaro FD, Valdovinos‐Ayala J, Percolla MI, Venturas MD, Pratt RB, Jacobsen AL (2019) Wood structure and function change with maturity: Age of the vascular cambium is associated with xylem changes in current‐year growth. Plant Cell Environ.42: 1816– 1831. doi: https://doi.org/10.1111/pce.13528
Sakai A, Larcher W (1987) Frost survival of plants. Ecol Stud. 62: 1– 321. doi: https://doi.org/10.1007/978-3-642-71745-1
| Interaction of drought and frost in tree ecophysiology: rethinking the timing of risks | Guillaume Charrier, Nicolas Martin-Stpaul, Claire Damesin, Nicolas Delpierre, Heikki Hänninen, José Torres-Ruiz, Hendrik Davi | <p>In temperate, boreal and alpine areas, the edges of plant distribution are strongly affected by abiotic constraints. For example, heat waves and drought are major constraints at low latitude and elevation while cold and frost are key factors at... |  | Tree biology and physiology | Rosana López | | 2020-04-28 21:07:27 | View |





